

(Albert, 2010)
A primeira etapa do desenvolvimento é a formação de um novo indivíduo originário da fusão de um gameta masculino haplóide, o espermatozóide, com um gameta feminino também haplóide, o ovócito, restaurando assim a diploidia na espécie humana. Desta maneira, fica assegurada a perenidade da espécie com descendentes diferentes de seus pais, mas portadores da mesma quantidade de material genético.
A fecundação é um processo fundamental nos organismos que possuem reprodução sexuada. Existe uma complexa inter-relação entre ambos os gametas. A presença do ovócito é fundamental na aquisição, antes da fertilização, de propriedades funcionais por parte do espermatozóide. Mesmo assim, este é imprescindível para a ativação do metabolismo celular do ovócito, o qual será encarregado de conduzir o desenvolvimento durante os primeiros momentos após a fecundação. Como resultado da fecundação, dois padrões serão definidos em princípio: o complemento cromossômico diplóide (2n) e o sexo cromossômico (XX, fêmea, e XY, macho). (Catala, 2003; Dumm, 2006)
PREPARAÇÃO DOS OVÓCITOS PARA A FECUNDAÇÃO
Durante a ovulação, ovócitos mamíferos são liberados do ovário para dentro da cavidade peritoneal próximo a entrada do oviduto, para dentro do qual são levados rapidamente. Eles estão cobertos por várias camadas de células foliculares (ou da granulosa) embebidas em uma matriz extracelular rica em hialuronana (ácido hialurônico). Essas células podem ajudar o ovócito a ser recolhido para dentro do oviduto, e elas também podem secretar sinais químicos (quimiotaxia) não identificados que atraem os espermatozóides para o ovócito.

PREPARAÇÃO DOS ESPERMATOZÓIDES PARA A FECUNDAÇÃO
Dois eventos devem ocorrer antes da fecundação: (1) a maturação dos espermatozóides no epidídimo, e (2) a capacitação dos espermatozóides no trato genital feminino.

(Moore, 2012)
EVENTOS MOLECULARES DA FECUNDAÇÃO
É uma complexa sequência de eventos moleculares coordenados que se inicia com o contato entre um espermatozóide e um ovócito e termina com a mistura dos cromossomos maternos e paternos, resultando no zigoto, um embrião unicelular. O processo de fecundação leva cerca de 24 horas, e ocorre normalmente na ampola da tuba uterina.
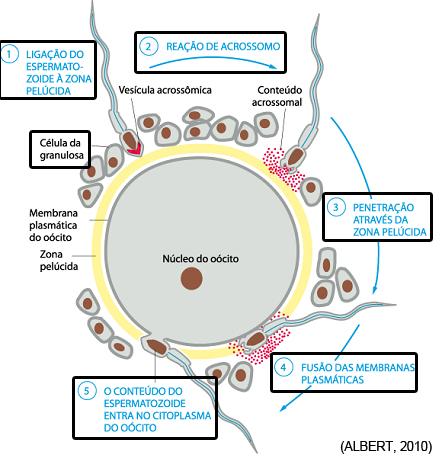
- Passagem do espermatozóide pela corona radiata
- Ligação do espermatozóide à zona pelúcida
- Reação de acrossomo (reação acrossômica)
- Penetração através da zona pelúcida
- Fusão das membranas plasmáticas
- O conteúdo do espermatozóide entra no citoplasma do ovócito
Passagem do espermatozóide pela corona radiata
A dispersão das células foliculares (ou da granulosa) da corona radiata que circunda o ovócito e a zona pelúcida parece resultar principalmente da ação da hialuronidase liberada pelo acrossoma do espermatozóide. As enzimas da mucosa tubárias também parecem auxiliar nesta dispersão, assim como os movimentos da cauda dos espermatozóides são importantes para a sua penetração na corona radiata.
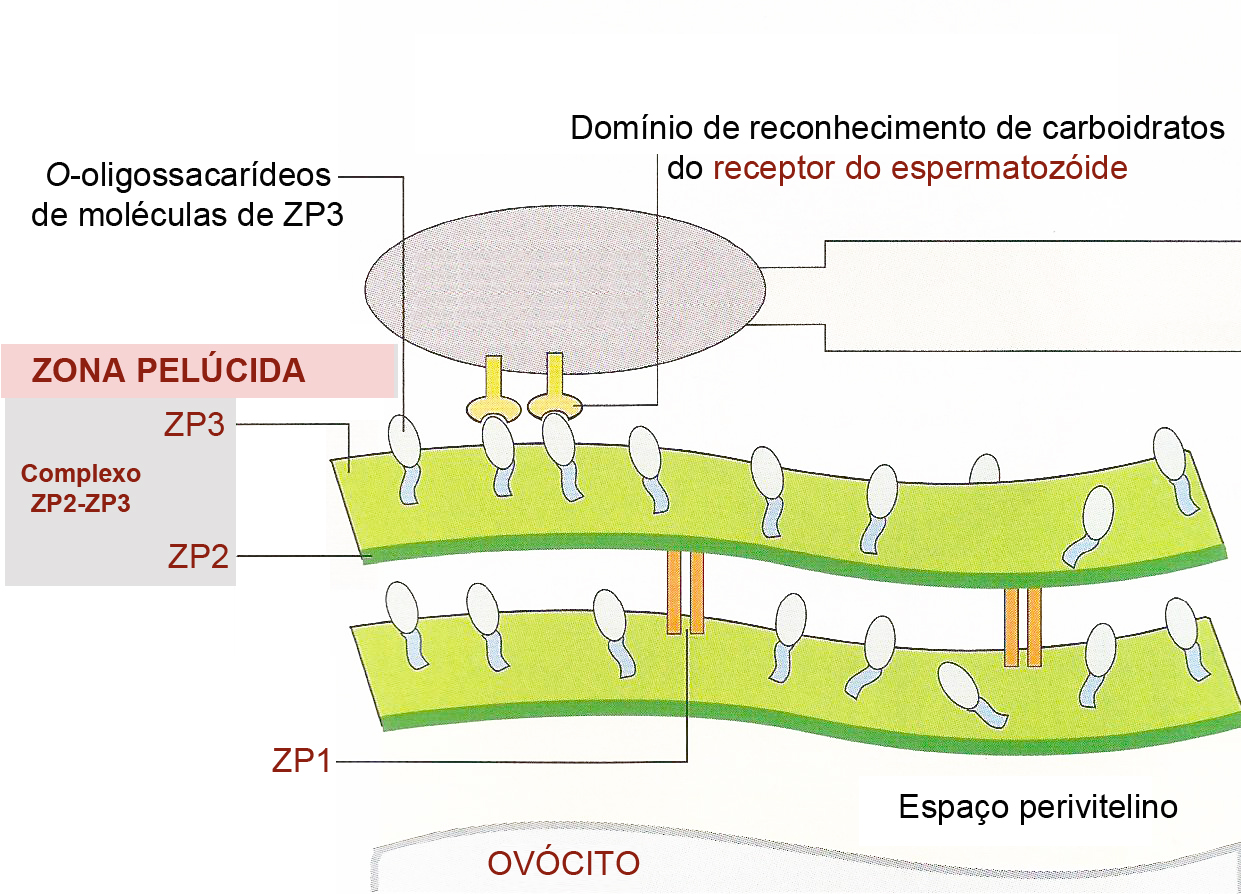
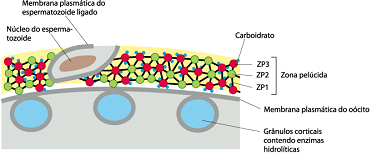
Ligação do espermatozóide à zona pelúcida
Durante a ligação com o espermatozóide, O-oligossacarídeos ligados à ZP3 interagem com os receptores de espermatozóides parecendo serem responsáveis, pelo menos em parte, pela ligação espécie-específica do espermatozóide à zona pelúcida. Entretanto, a ligação do espermatozóide à zona pelúcida é complexa e envolve tanto mecanismos dependentes quanto independentes de ZP3, assim como uma variedade de proteínas da superfície do espermatozóide. A zona pelúcida induz o espermatozóide a sofrer a reação do acrossoma ou reação acrossômica.
(Kierszenbaum, 2008)
Reação do acrossomo ou reação acrossômica

(Kierszenbaum, 2008)
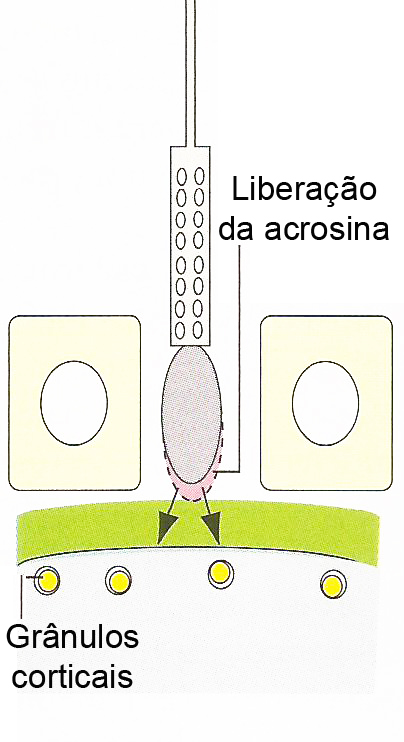
A reação acrossômica é mediada por um aumento na concentração de íons Ca2+ no interior do espermatozóide, através da abertura de canais de Ca2+ na sua superfície. Esse fluxo de Ca2+ é acompanhado por um ingresso de Na+ e pela saída de H+, o que gera um aumento do pH intracelular do gameta. Esta abertura dos canais seria facilitada por uma alteração no potencial de membrana produzido durante o processo de capacitação. A soma destes fatores leva a fusão da membrana acrossômica externa com a membrana plasmática do espermatozóide, formando-se vesículas e poros, e possibilitando a liberação de enzimas para facilitar a passagem deste gameta pela zona pelúcida. Finalizada a reação, a membrana acrossômica interna passa a constituir a membrana externa da cabeça do espermatozóide, em continuidade com o resto de sua membrana plasmática. A primeira, então, é a que entra em contato com a zona pelúcida. Durante essa reação, são liberadas inúmeras enzimas, no entanto, a acrosina, que se encontra unida à membrana acrossômica interna, seria a mais importante na construção do caminho do espermatozóide através da zona pelúcida. Alguns autores sustentam que a acrosina não seria liberada ao meio e, sim, manter-se-ia unida à membrana acrossômica interna durante todo o trajeto.
Penetração através da zona pelúcida
Somente os espermatozóides que sofrerem a reação acrossômica é que podem penetrar na zona pelúcida. O primeiro espermatozóide a chegar na zona pelúcida se liga à ZP3, um dos três componentes glicoproteicos. A ligação à ZP3 induz a liberação da acrosina pela membrana acrossômica interna, facilitando a penetração da cabeça do espermatozóide na zona pelúcida.
(Kierszenbaum, 2008)
(Kierszenbaum, 2008
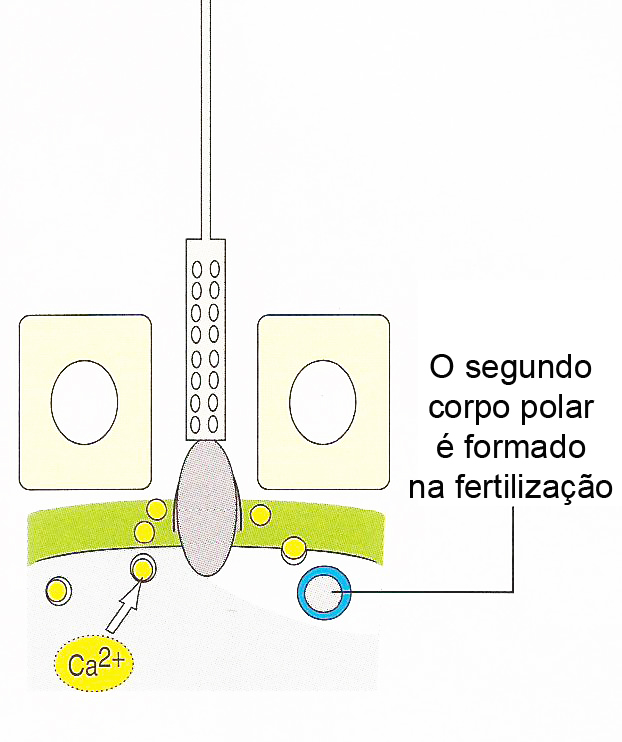
O primeiro espermatozóide a penetrar na zona pelúcida se funde com a membrana do ovócito e induz a exocitose Ca2+ -dependente dos grânulos corticais localizados logo abaixo da membrana plasmática, em um processo chamado de reação cortical. O conteúdo liberado dos grânulos corticais inativa a ZP3, de modo que ela não pode mais ligar-se a membrana do espermatozóide. Há também a clivagem da ZP2, endurecendo a zona pelúcida de maneira que os espermatozóides não podem penetrá-la. Juntas, estas alterações promovem a reação da zona resultando em um bloqueio à poliespermia.
(Albert, 2010)
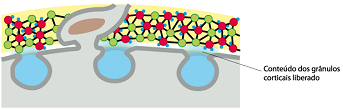
Fusão das membranas plasmáticas do espermatozóide com a do ovócito secundário
(Kierszenbaum, 2008)
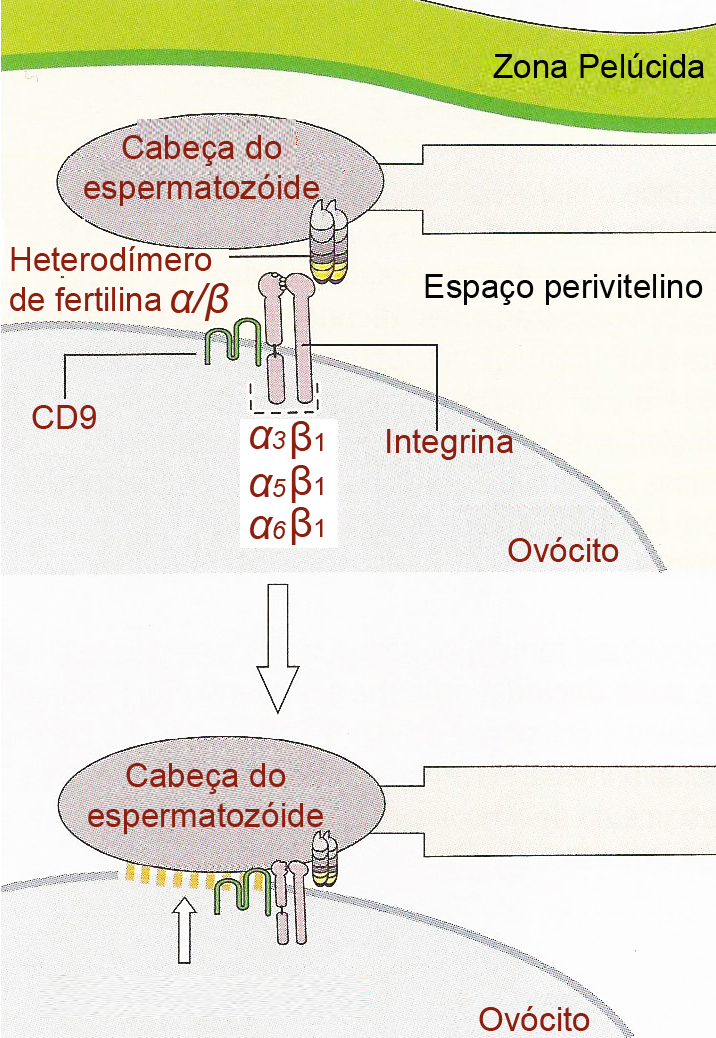
Após a passagem pela zona pelúcida, o espermatozóide liga-se à membrana plasmática do ovócito, inclinando-se sobre as microvilosidades na superfície do gameta feminino. Rapidamente as microvilosidades vizinhas se agrupam em torno do espermatozóide para assegurar que este seja aderido de forma firme. Os mecanismos moleculares responsáveis pela fusão entre as membranas dos dois gametas são em grande parte desconhecidos, embora, após um número de ativações artificiais, duas proteínas de membrana têm sido apontadas como fundamentais para a fusão. Uma é a proteína transmembrana da superfamília das imunoglobulinas específica do espermatozóide chamada de Izumo, que torna-se exposta durante a reação acrossômica. A única proteína na superfície do ovócito que se demonstrou necessária foi a proteína CD9, que é um membro da família das tetraspaninas, assim chamadas pois possuem quatro segmentos que transpõem a membrana. Na presença do complexo CD9-integrina e da fertilina α/β, as membranas plasmáticas do espermatozóide e do ovócito se fundem através de mecanismos de adesão intercelular.
Fusão da membranas plasmáticas – ativação do ovócito secundário
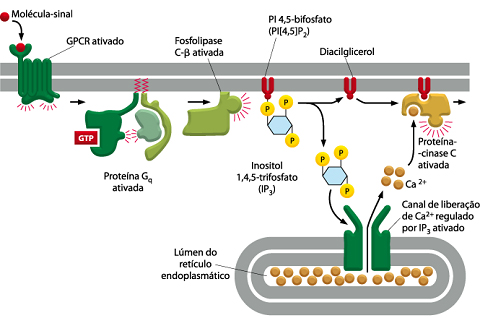
Quando o espermatozóide funde-se com a membrana plasmática de um ovócito de uma maneira normal, isso causa um aumento local de Ca2+ citosólico, que se difunde como uma onda através da célula. A onde se propaga por feedback positivo: a elevação do Ca2+ citosólico causa a abertura dos canais de Ca2+, permitindo que ainda mais Ca2+ entre no citosol. A onda inicial da liberação de Ca2+ normalmente é seguida, dentro de poucos minutos, por oscilações de Ca2+ que persistem por várias horas. O espermatozóide que realizou a fusão dispara a onda e as oscilações de Ca2+ por introduzir um fator dentro do citosol do ovócito, que causa um aumento na concentração de inositol 1,4,5-trifosfato (IP3), o qual libera Ca2+ do retículo endoplasmático e inicia a onda e as oscilações de Ca2+. Um candidato forte para ser este fator critico é uma fosfolipase C específica de espermatozóides, a PLC ζ que cliva diretamente o fosfoinositol 4,5-bifosfato (PI[4,5]P2) para produzir IP3 e diacilglicerol.
(Albert, 2010)
O ovócito termina sua segunda divisão meiótica imediatamente após a penetração do espermatozóide, formando um ovócito maduro e um segundo corpo polar. Os cromossomos maternos se descondensam e o núcleo do ovócito maduro torna-se o pronúcleo feminino.
O conteúdo do espermatozóide entra no citoplasma do ovócito
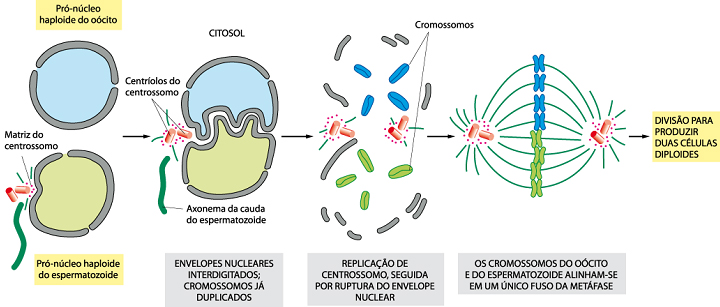
A cabeça e a cauda do espermatozóide entram no citoplasma do ovócito, mas a membrana plasmática e as mitocôndrias ficam para trás, e a cauda degenera. Dentro do citoplasma do ovócito, o núcleo do espermatozóide aumenta para formar o pronúcleo masculino. Morfologicamente, os dois pronúcleos são indistinguiveis. Durante o crescimento dos pronúcleos, eles replicam o seu DNA-1n (haplóide), 2c (duas cromátides). O ovócito contendo dois pronúcleos haplóides é chamado de oótide. Os pronúcleos se aproximam e seus envelopes nucleares formam interdigitações. Um centrossomo se forma em torno dos centríolos fornecidos pelo espermatozóide, uma vez que em ovócitos não fecundados, estas estruturas não estão presentes. O centrossomo se duplica, os envelopes nucleares se rompem, e finalmente, os cromossomos dos gametas são integrados em um fuso mitótico único, que organiza a primeira divisão mitótica da clivagem. Logo que os pronúcleos se fundem em uma agregação de cromossomos única e diplóide, a oótide torna-se um zigoto.
(Albert, 2010)